










Canada Warbler Cardellina canadensis Scientific name definitions
- LC Least Concern
- Names (41)
- Monotypic
Len R. Reitsma, Michael T. Hallworth, Marissa McMahon, and Courtney J. Conway
Version: 2.0 — Published May 7, 2020
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Catalan | bosquerola del Canadà |
| Czech | lesňáček kanadský |
| Danish | Canadasanger |
| Dutch | Canadese Zanger |
| English | Canada Warbler |
| English (United States) | Canada Warbler |
| French | Paruline du Canada |
| French (France) | Paruline du Canada |
| German | Kanadawaldsänger |
| Greek | Καναδέζικη Πάρουλα |
| Haitian Creole (Haiti) | Ti Tchit Kanada |
| Hebrew | סבכון קנדי |
| Hungarian | Örvös lombjáró |
| Icelandic | Haustskríkja |
| Japanese | クロボシアメリカムシクイ |
| Lithuanian | Kanadinė vilsonija |
| Norwegian | kanadaparula |
| Polish | wilsonka kanadyjska |
| Portuguese (Brazil) | mariquita-do-canadá |
| Portuguese (Portugal) | Mariquita-do-canadá |
| Romanian | Frunzar cu burtă galbenă |
| Russian | Канадская вильсония |
| Serbian | Kanadska cvrkutarka |
| Slovak | horárik kanadský |
| Slovenian | Apalaški frfotavček |
| Spanish | Reinita Canadiense |
| Spanish (Chile) | Reinita de Canadá |
| Spanish (Costa Rica) | Reinita Pechirrayada |
| Spanish (Cuba) | Bijirita del Canadá |
| Spanish (Dominican Republic) | Cigüita del Canadá |
| Spanish (Ecuador) | Reinita Collareja (de Canadá) |
| Spanish (Honduras) | Chipe Collarejo |
| Spanish (Mexico) | Chipe de Collar |
| Spanish (Panama) | Reinita Collareja |
| Spanish (Peru) | Reinita de Canada |
| Spanish (Puerto Rico) | Reinita de Canadá |
| Spanish (Spain) | Reinita canadiense |
| Spanish (Venezuela) | Reinita Canadiense |
| Swedish | kanadaskogssångare |
| Turkish | Kanada Ötleğeni |
| Ukrainian | Болотянка строкатовола |
Cardellina canadensis (Linnaeus, 1766)
PROTONYM:
Muscicapa canadensis
Linnaeus, 1766. Systema Naturæ. Editio duodecima reformata. Tomus I [part 1], p.327.
TYPE LOCALITY:
Canada.
SOURCE:
Avibase, 2023
Definitions
- CARDELLINA
- canadensis
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
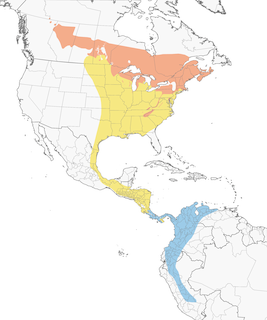
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Canada Warbler















