












Pacific Golden-Plover Pluvialis fulva Scientific name definitions
- LC Least Concern
- Names (56)
- Monotypic
Oscar W. Johnson, Peter G. Connors, and Peter Pyle
Version: 1.1 — Published April 15, 2021
Revision Notes
Revision Notes
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Asiatiese Goue Strandkiewiet |
| Albanian | Gjelaci siberian ngjyrëartë |
| Arabic | زقزاق ذهبي سيبيري |
| Asturian | Pilordu dorñu siberianu |
| Azerbaijani | Asiya qonurqanadı |
| Basque | Urre-txirri siberiarra |
| Bulgarian | Сибирска булка |
| Catalan | daurada del Pacífic |
| Chinese | 太平洋金斑鴴 |
| Chinese (SIM) | 金斑鸻 |
| Croatian | mali zlatar |
| Czech | kulík pacifický |
| Danish | Sibirisk Hjejle |
| Dutch | Aziatische Goudplevier |
| English | Pacific Golden-Plover |
| English (HAW) | Kōlea - Pacific Golden-Plover |
| English (UK) | Pacific Golden Plover |
| English (United Arab Emirates) | Pacific Golden Plover |
| English (United States) | Pacific Golden-Plover |
| Finnish | siperiankurmitsa |
| French | Pluvier fauve |
| French (France) | Pluvier fauve |
| Galician | Píllara dourada siberiana |
| German | Tundra-Goldregenpfeifer |
| Greek | Βροχοπούλι του Ειρηνικού |
| Hebrew | חופזי קטן |
| Hungarian | Ázsiai pettyeslile |
| Icelandic | Glitlóa |
| Indonesian | Cerek kernyut |
| Italian | Piviere orientale |
| Japanese | ムナグロ |
| Korean | 검은가슴물떼새 |
| Latvian | Tundras tārtiņš |
| Lithuanian | Tundrinis sėjikas |
| Malayalam | പൊൻമണൽക്കോഴി |
| Mongolian | Азийн сүвээ цагаан |
| Norwegian | sibirlo |
| Odia | ଘଣ୍ଟୁଆଳ |
| Persian | سلیم طلایی خاوری |
| Polish | siewka złotawa |
| Portuguese (Portugal) | Tarambola-dourada-siberiana |
| Romanian | Ploier auriu asiatic |
| Russian | Бурокрылая ржанка |
| Serbian | Tihookeanski zlatni vivak |
| Slovak | kulík krátkochvostý |
| Slovenian | Sibirska zlata prosenka |
| Spanish | Chorlito Dorado Siberiano |
| Spanish (Chile) | Chorlo dorado del Pacífico |
| Spanish (Costa Rica) | Chorlito Dorado del Pacífico |
| Spanish (Ecuador) | Chorlo Dorado del Pacífico |
| Spanish (Mexico) | Chorlo Dorado del Pacífico |
| Spanish (Spain) | Chorlito dorado siberiano |
| Swedish | sibirisk tundrapipare |
| Thai | นกหัวโตหลังจุดสีทอง |
| Turkish | Küçük Altın Yağmurcun |
| Ukrainian | Сивка бурокрила |
Revision Notes
In this partial revision, Oscar W. Johnson updated the Identification, Timing and Routes of Migration, Effects of Human Activity, and Other, with additional new references added to each of these articles.
Pluvialis fulva (Gmelin, 1789)
PROTONYM:
Charadrius fulvus
Gmelin, 1789. Systema Naturae. Editio decima tertia, aucta, reformata. Cura Jo. Frid. Gmelin (etc.). Tomus I. Pars II 1 pt2, p.687.
TYPE LOCALITY:
Tahiti.
SOURCE:
Avibase, 2023
Definitions
- PLUVIALIS
- pluvialis
- fulva
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
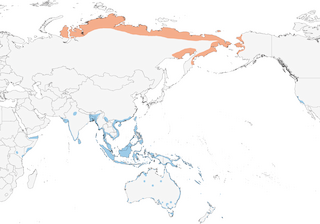
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Pacific Golden-Plover















