











Purple Gallinule Porphyrio martinica Scientific name definitions
- LC Least Concern
- Names (48)
- Monotypic
Richard L. West and Gene K. Hess
Version: 1.0 — Published March 4, 2020
Text last updated January 1, 2002
Text last updated January 1, 2002
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Amerikaanse Koningriethaan |
| Bulgarian | Американска султанка |
| Catalan | polla blava americana |
| Czech | slípka žlutonohá |
| Danish | Amerikansk Sultanhøne |
| Dutch | Amerikaans Purperhoen |
| English | Purple Gallinule |
| English (South Africa) | Purple Gallinule (American) |
| English (United States) | Purple Gallinule |
| French | Talève violacée |
| French (France) | Talève violacée |
| German | Zwergsultanshuhn |
| Greek | Αμερικανική Πορφυροπουλάδα |
| Haitian Creole (Haiti) | Poul sultan |
| Hebrew | פורפיריה אמריקנית |
| Hungarian | Amerikai szultántyúk |
| Icelandic | Flóðhæna |
| Italian | Pollo sultano della Martinica |
| Japanese | アメリカムラサキバン |
| Lithuanian | Amerikinė sultonvištė |
| Norwegian | amerikapurpurhøne |
| Polish | sułtanka amerykańska |
| Portuguese (Brazil) | frango-d'água-azul |
| Portuguese (Portugal) | Camão-americano |
| Romanian | Găinușă americană |
| Russian | Малая султанка |
| Serbian | Američka sultanka |
| Slovak | sultánka indigová |
| Slovenian | Ameriška sultanka |
| Spanish | Calamoncillo Americano |
| Spanish (Argentina) | Pollona Azul |
| Spanish (Chile) | Tagüita purpúrea |
| Spanish (Costa Rica) | Gallareta Morada |
| Spanish (Cuba) | Gallareta azul |
| Spanish (Dominican Republic) | Gallareta Azul |
| Spanish (Ecuador) | Gallareta Púrpura |
| Spanish (Honduras) | Gallineta Púrpura |
| Spanish (Mexico) | Gallineta Morada |
| Spanish (Panama) | Gallareta Morada |
| Spanish (Paraguay) | Polla azul |
| Spanish (Peru) | Polla de Agua Morada |
| Spanish (Puerto Rico) | Gallareta Azul |
| Spanish (Spain) | Calamoncillo americano |
| Spanish (Uruguay) | Polla Azul |
| Spanish (Venezuela) | Gallito Azul |
| Swedish | amerikansk sultanhöna |
| Turkish | Sarı Bacaklı Sazhorozu |
| Ukrainian | Султанка американська |
Porphyrio martinica (Linnaeus, 1766)
PROTONYM:
Fulica martinica
Linnaeus, 1766. Systema Naturæ. Editio duodecima reformata. Tomus I [part 1], p.259.
TYPE LOCALITY:
Martinique, West Indies.
SOURCE:
Avibase, 2023
Definitions
- PORPHYRIO
- porphyrio
- martinica / martinicana / martinicensis / martinicus
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
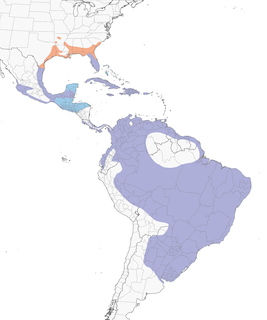
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Purple Gallinule















