

























Red-tailed Hawk Buteo jamaicensis Scientific name definitions
- LC Least Concern
- Names (38)
- Subspecies (14)
C. R. Preston and R. D. Beane
Version: 1.0 — Published March 4, 2020
Text last updated May 20, 2009
Version: 1.0 — Published March 4, 2020
Text last updated May 20, 2009
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Asturian | Buzacu raurroxu |
| Basque | Zapelatz buztangorria |
| Bulgarian | Червеноопашат мишелов |
| Catalan | aligot cua-roig |
| Croatian | crvnorepi škanjac |
| Czech | káně rudochvostá |
| Dutch | Roodstaartbuizerd |
| English | Red-tailed Hawk |
| English (United States) | Red-tailed Hawk |
| Finnish | amerikanhiirihaukka |
| French | Buse à queue rousse |
| French (France) | Buse à queue rousse |
| Galician | Miñato rabirrubio |
| German | Rotschwanzbussard |
| Greek | Αμερικανική Γερακίνα |
| Haitian Creole (Haiti) | Malfini ke wouj |
| Icelandic | Ryðvákur |
| Italian | Poiana codarossa |
| Japanese | アカオノスリ |
| Norwegian | rødhalevåk |
| Polish | myszołów rdzawosterny |
| Russian | Краснохвостый канюк |
| Serbian | Crvenorepi mišar |
| Slovak | myšiak hrdzavochvostý |
| Slovenian | Rdečerepa kanja |
| Spanish | Busardo Colirrojo |
| Spanish (Costa Rica) | Gavilán Colirrojo |
| Spanish (Cuba) | Gavilán de monte |
| Spanish (Dominican Republic) | Guaraguao |
| Spanish (Honduras) | Gavilán Cola Roja |
| Spanish (Mexico) | Aguililla Cola Roja |
| Spanish (Panama) | Gavilán Colirrojo |
| Spanish (Puerto Rico) | Guaraguao Colirrojo |
| Spanish (Spain) | Busardo colirrojo |
| Spanish (Venezuela) | Gavilán Colirrojo |
| Swedish | rödstjärtad vråk |
| Turkish | Kızıl Kuyruklu Şahin |
| Ukrainian | Канюк неоарктичний |
Buteo jamaicensis (Gmelin, 1788)
PROTONYM:
Falco jamaicensis
Gmelin, 1788. Systema Naturae. Editio decima tertia, aucta, reformata. Cura Jo. Frid. Gmelin (etc.). Tomus I [pars I], p.266.
TYPE LOCALITY:
Jamaica.
SOURCE:
Avibase, 2023
Definitions
- BUTEO
- buteo
- jamaica / jamaicae / jamaicana / jamaicensis
The Key to Scientific Names
Legend Overview
UPPERCASE: current genus
Uppercase first letter: generic synonym
● and ● See: generic homonyms
lowercase: species and subspecies
●: early names, variants, misspellings
‡: extinct
†: type species
Gr.: ancient Greek
L.: Latin
<: derived from
syn: synonym of
/: separates historical and modern geographic names
ex: based on
TL: type locality
OD: original diagnosis (genus) or original description (species)
Account navigation Account navigation
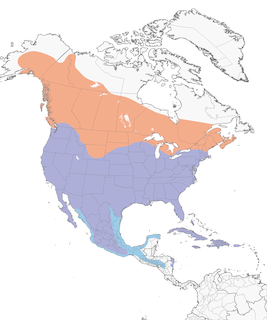
- Year-round
- Migration
- Breeding
- Non-Breeding
Distribution of the Red-tailed Hawk